
{kind=link}
Исследование, опубликованное в журнале Science и проведенное под руководством Кембриджского университета и Хельсинкского университета, раскрывает регуляторные механизмы, управляющие формированием ксилемы, и предлагает новые сведения о том, как растения строят как структурные, так и запасающие ткани.
В процессе эволюции у растений сформировалось множество сигнальных молекул, которые, работая вместе, создают архитектуру сосудистой системы, охватывающую весь их организм.
Развитие сосудистой системы тщательно регулируется путем настройки сигнальных воздействий на различных этапах, что помогает определить, будет ли растение производить больше водопроводящих клеток или больше запасающих клеток.
Растения могут переключаться между этими приоритетами роста, поскольку их водопроводящая ксилемная ткань состоит из двух различных типов клеток:
- Клетки сосудов с утолщенными вторичными клеточными стенками, обогащенными лигнином, которые обеспечивают быстрое движение воды
- Паренхимные клетки с тонкими первичными клеточными стенками, которые запасают воду и питательные вещества, такие как крахмал.
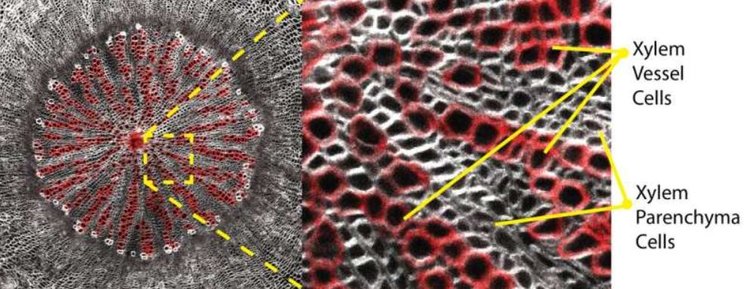
Чтобы выяснить, как растения тонко регулируют баланс между этими типами клеток, исследователи из лабораторий профессора Ика Хелариутты в университетах Кембридже и Хельсинки изучили однокопийный генный мутант модельного растения Arabidopsis thaliana, называемый overachiever (ovac), который производит избыток сосудистых клеток за счет паренхимных клеток.
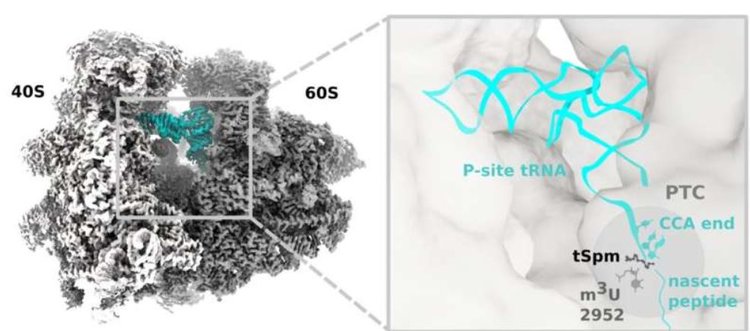
Команда исследователей установила, что ген OVAC представляет собой метилтрансферазу рРНК, ответственную за модификацию основания m3U2952 именно в пептидилтрансферазном центре 25S рРНК у Arabidopsis.
Затем они сосредоточились на термоспермине, небольшой положительно заряженной молекуле полиамина, которая, как известно, регулирует дифференцировку сосудов, увеличивая трансляцию транскрипционных факторов SAC51, которые, в свою очередь, ингибируют инициацию ксилемных сосудов.
«Было неясно, как работает эта трансляционная регуляция и как она корректируется, — сказал соавтор, доктор Донхви Ко из лаборатории Сейнсбери Кембриджского университета. — Мы также знали, что полиамины взаимодействуют с рибосомами, связываясь с рибосомальной РНК (рРНК), поэтому мы хотели изучить и этот аспект».
Рибосомы как сенсоры сигналов
Исследователи обнаружили, что термоспермин связывается с метилированными рибосомами, позволяя рибосоме действовать в качестве сигнального сенсора, способствующего трансляции SAC51.
Рибосомы — это белковые фабрики, и они часто взаимодействуют и связываются с полиаминами, такими как термоспермин.
Однако, несмотря на роль связанных с рибосомами полиаминов в разнообразных биологических процессах, включая регуляцию пролиферации, дифференцировки, аутофагии и старения клеток, до сих пор не было выявлено каких-либо специфических клеточных функций, связанных с ними.
Исследователи из лаборатории профессора Алана Уоррена в Кембриджском институте медицинских исследований и лаборатории доцента Финна Кирпекара в Университете Южной Дании провели углубленное исследование химических модификаций и структурной биологии рибосомы Arabidopsis, чтобы визуализировать происходящие процессы.
«Метиляция происходит в пептидилтрансферазном центре рибосомы, где может катализироваться образование пептидной связи. Эта модификация позволяет термоспермину связываться в том же месте, соединяя ключевые компоненты регуляции трансляции», — сказал соавтор, доктор Александр Файль из Кембриджского института медицинских исследований.
В результате дальнейших исследований было обнаружено, что связывание термоспермина с метилированными рибосомами влияет на второй фактор транскрипции, регулирующий развитие сосудов, называемый LHW, который усиливает инициацию образования сосудов. Термоспермин ингибировал LHW.
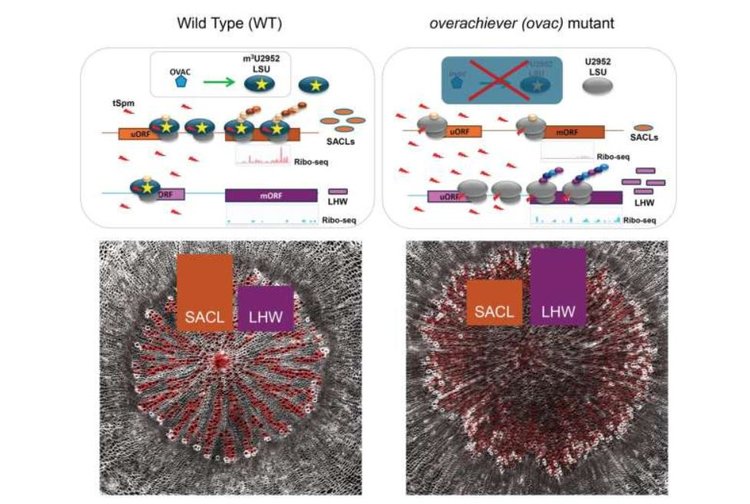
«Когда трансляция SAC51 и LHW регулируется термоспермином в растениях дикого типа, устанавливается нормальный баланс между клетками сосудов и паренхимы», — сказала соавтор, доктор Ева Хеллманн из лаборатории Сейнсбери Кембриджского университета.
Однако у мутанта термоспермин не может стабильно связываться с неметилированными рибосомами, что препятствует этой регуляции и позволяет осуществлять высокий уровень трансляции LHW, что нарушает нормальное формирование сосудистой сети.
Исследование показало, что термоспермин регулирует оба транскрипционных фактора по-разному. Он является бифункциональным регулятором трансляции для SAC51 и LHW, но для его бифункциональной регуляции требуется специфическое метилирование, m3U2952. Он действует, стимулируя трансляцию SACL и ингибируя трансляцию LHW.
«Это исследование проливает свет на то, как растения точно настраивают развитие сосудистой системы , чтобы определить судьбу своих сосудистых клеток», — сказала соавтор, доктор Райли Руонала из Хельсинкского университета. «Эти результаты потенциально могут повлиять на характеристики растений, начиная от засухоустойчивости и заканчивая ростом корней/клубней у сельскохозяйственных культур, а также формированием древесины», подчеркнула она.
Хотя исследование проводилось на модельном растении арабидопсис (резуховидка Таля), оно указывает на то, что аналогичные сигналы могут происходить и в других растениях. Например, у деревьев эти сигналы потенциально могут быть настроены на образование большого количества водопроводящих сосудов для поддержания высокого роста. У редиса те же сигналы могут быть скорректированы таким образом, чтобы способствовать образованию запасающих клеток в корне, позволяя растению накапливать больше энергии.
Источник: Источник: University of Cambridge.
На заглавном фото: поперечный срез гипокотиля 8-недельного растения Arabidopsis thaliana. Мутант ovac демонстрирует усиленную вторичную дифференциацию сосудов (красный цвет) за счет паренхимных клеток. На изображениях A2 и B2 показаны только клетки сосудов. Окрашивание SR2200 и фуксином. Автор фото: Донхви Ко.
