Жгутики – это необязательный структурный компонент бактериальной клетки. Они могут быть удалены без нарушения метаболизма клетки. Первые сведения о наличии жгутиков у бактерии сообщил в 1838 году немецкий естествоиспытатель Г. Эренбер. В 1897 году немецкий ботаник В. Мигула дал их морфологическое описание. Жгутики у бактерий видны только в электронном микроскопе. В световом микроскопе, без специальной обработки отдельные жгутики увидеть нельзя. В процессе окрашивания бактерий по Граму жгутики не видны.
{kind=link}
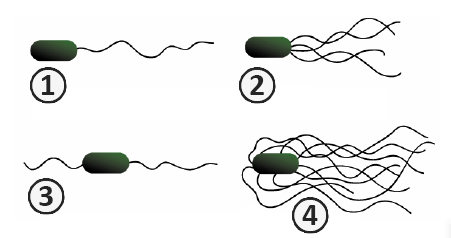
Типы бактерий в зависимости от расположения и числа жгутиков
В зависимости от расположения и числа жгутиков на поверхности клетки различают следующие типы бактерий:
- монотрихи – имеют только один жгутик (род Caulobacter и род Vibrio);
- лофотрихи – имеют на одном или на обоих полюсах клетки пучок жгутиков (род Pseudomonas);
- амфитрихи – имеют по одному жгутику на обоих полюсах клетки (род Spirillum);
- перитрихи – имеется большое количество жгутиков, располагающихся по всей поверхности клетки (род Erwinia).
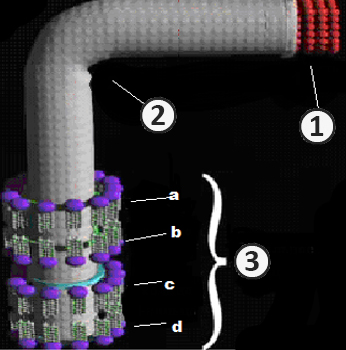
Структура жгутика
Электронно-микроскопические исследования выявляют сложную структурную организацию жгутиков.
Жгутик состоит из трех частей: нити, крюка, базального тельца.
Жгутик закреплен в цитоплазматической мембране и клеточной стенке с помощью базального тельца. В структуру последнего входит стержень и кольца.
Количество колец базального тельца у грамотрицательных и грамположительных бактерий различно.
У грамотрицательных бактерий – четыре кольца:L, P, S, M.
L и P – наружная пара колец. S и M – внутренняя пара колец.
Кольца жгутика грамотрицательных бактерий закреплены в разных местах:
- L-кольцо – в наружной мембране клеточной стенки;
- P-кольцо – в пептидогликановом слое клеточной стенки;
- S-кольцо – в периплазматическом пространстве клеточной стенки;
- M-кольцо – в цитоплазматической мембране.
Жгутики грамположительных бактерии характеризуются базальным тельцем более простого строения. В данном случае оно состоит только из внутренней пары колец – S и M, размещенных в цитоплазматической мембране и клеточной стенке.
{kind=link}
Химический состав жгутиков
Химический состав жгутиков однообразен. Они состоят из белка флагеллина (от латинского «flagellum» – жгутик) с молекулярной массой 25000–60000. В аминокислотном составе данного белка преобладают глутаминовая и аспарагиновая аминокислоты. Количество ароматических аминокислот в флагеллине незначительно. Триптофан, цистеин и цистин содержаться в следовых количествах или совсем отсутствуют.
Движение бактерий при помощи жгутиков
Как указывалось ранее, жгутики являются органами движения бактерий. Характер движения определяется особенностью расположения жгутиков:
Скорость движения бактерий при помощи жгутиков различна. Большинство подвижных форм бактерий за одну секунду проходят расстояние, близкое размерам их тел. Самой подвижной бактерией считается холерный вибрион. При длине тела в 2 мкм, он проходит за одну секунду до 30 мкм.
Работа бактериального жгутика подобна работе корабельного винта. Если у клетки много жгутиков, то при передвижении они собираются в пучок, образующий своеобразный пропеллер. Пучок жгутиков быстро вращается против часовой стрелки, создавая силу, заставляющую бактерию двигаться почти прямолинейно. После того, как направление вращения жгутиков изменяется, пучок жгутиков расплетается, клетка останавливается и начинает хаотически вращаться и менять ориентацию. В момент, когда все жгутики снова начнут вращаться синхронно против часовой стрелки, образуя пропеллер, направление поступательного движения бактериальной клетки изменится.
Поскольку у грамположительных бактерий отсутствует наружная пара колец, то считается, что для вращения жгутиков достаточно только внутренней пары колец (S и M). Данные кольца, соединенные с вращающимся стержнем, выступающим наружу, образуют своеобразный электромотор, обеспечивающий движение жгутика.
В качестве источника энергии для вращения жгутиков используется протодвижущая сила, возникающая в цитоплазматической мембране. Происходит это следующим образом. На внешней стороне (периферии) кольца M расположены белки MotB. В участок цитоплазматической мембраны, примыкающей к краям колец M и S, встроены белки MotA.
Вращающий момент возникает за счет взаимодействия субъединиц белка MotB с белковыми субъединицами MotA. В белковых субъединицах MotA имеются два протонных полуканала. Через них переносятся протоны из периплазматического пространства клеточной стенки в цитоплазму бактерий. В результате переноса протонов через белки MotА и MotВ происходит вращение кольца М. Один полный оборот данного кольца происходит при переносе через мембрану около 1000 протонов.